Conduction of Action Potentials along Axons
A typical neuron is comprised of several distinct subcellular compartments (Figure 1):
- cell body (soma)
- dendrites (dendritic tree)
- initial segment (axon hillock)
- axon
- nerve terminal
The neuron is functionally polarized. Electrical information passes from the presynaptic nerve terminal to the dendrites or cell body of the postsynaptic neuron and then travels down the axon to the nerve terminal, where it is transmitted to the next neuron. In general, there is a one-way flow of electrical excitation from the presynaptic neuron to the postsynaptic neuron.
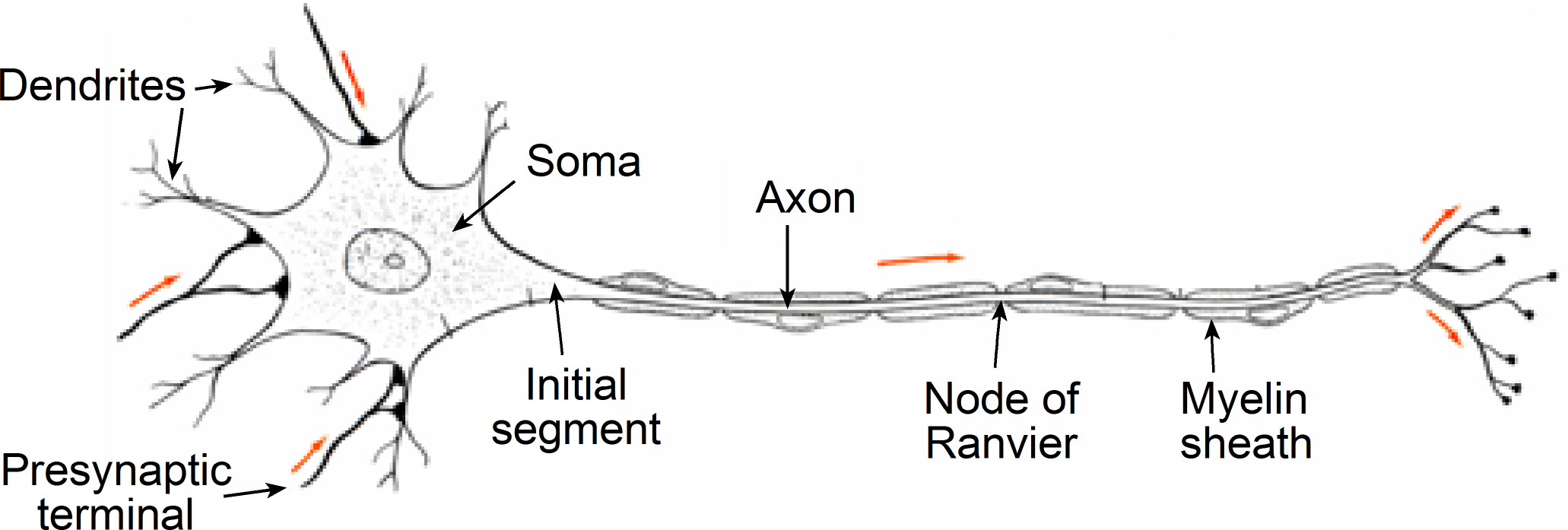
Figure 1 Structure of a typical neuron. Arrows indicate the normal direction of travel of electrical excitation.
The action potential typically initiates in the initial segment, since this is a region where there is a high concentration of Na+ channels, and then propagates down the axon by depolarizing the neighboring axon membrane. This self-propagating wave of depolarization travels down the axon and ends in the nerve terminal.
Passive Cable Properties
The passive electrical properties of the axon membrane are more complicated than those of a simple spherical cell described previously. They can be described using a similar mathematical model to that originally first used to analyze the electrical properties of transatlantic telegraph cables. As a consequence, the passive electrical properties of axons are known as its cable properties.
If a current is injected into the center of an axon at a single point the current can do one of two things (Figure 2). It can flow axially along the interior of the axon or it can flow back to ground across the membrane. The relative amount of current that crosses the membrane versus the amount of current that flows axially depends upon the resistance of the membrane relative to the resistance of the axial current path. With each increment in distance along the axon, the fraction of the injected current that flows down the axon again faces two paths and the amount of current flowing down the nerve fiber will gradually decrement as will the amount of current crossing the membrane resistance. As a consequence, the injected current will have a steadily decreasing influence on the membrane potential with increasing distance from the site of current injection.
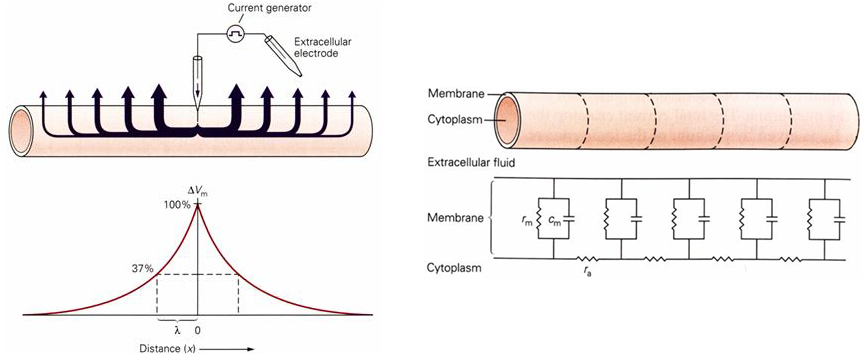
Figure 2 (left panel) (upper figure) Current flow in an axon following current injection at a single point in the axon. (lower figure) Change in membrane voltage as a function of distance from the point of current injection. (right panel) Electrical model of the axon as a series of RC circuits.
The axon cable can be mathematically modeled as a series of RC circuits connected together (Figure 2). The cable is imagined to be cut-up into small segments, with each segment corresponding to an RC circuit (with membrane resistance \(r_{m}\)) that is linked to its neighbors by an access resistance (\(r_{a}\)).
A measure of the electrical performance of a cable is known as the length constant (\(\lambda\)).
\[\lambda = \sqrt{\frac{r_{m}}{r_{a}}}\ \ \ \ \ \ \ \ \ \ \ \ \ \ \ \ \ (1)\]
The length constant is the distance over which the steady-state membrane potential drops to 37% of the original amplitude. The conduction properties of the cable are best when \(r_{a}\ \)is relatively small and \(r_{m}\) is relatively large. In other words, decreasing the axial resistance increases the length constant and increasing the membrane resistance, increases the length constant.
Axons are not very good conductors by the standards of typical electrical components and length constants are relatively short. In a typical axon the voltage will decrement by about 95% within 1-2 mm of the site of current injection.
Local Circuit Currents
The key to understanding how an action potential propagates down an axon is to understand local circuit flow (Figure 3). The artificial injection of a current at one point in an axon causes currents to flow for some distance from the point of current injection. These are known as local circuit currents and the distance over which these currents flow is determined by the length constant. Local circuit current flow acts to depolarize the cell membrane for some distance from the site of current injection.
Current flow through the axon surrounding the peak of an action potential is directly analogous to the injected current model (Figure 3). At the peak of the action potential there is an inward flow of current carried by sodium ions. The membrane on either side of the action potential peak will be depolarized by local circuit flow, in the same way as for the artificially injected current. The region of membrane in front of the action potential wave front, whose membrane potential is pushed above the threshold potential by local circuit flow, will initiate an action potential, which is now some distance further down the axon from the original action potential. This cycle of local circuit flows initiating an action potential in front of the traveling action potential wave front repeats itself continuously as the action potential travels down the axon.
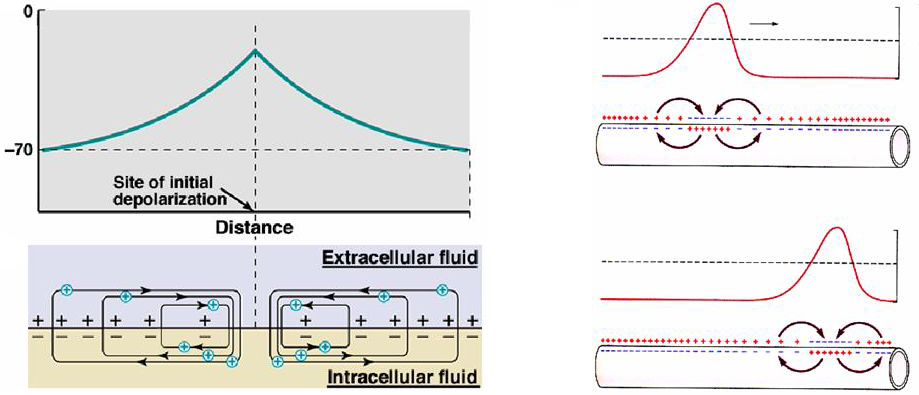
Figure 3 (left panel) Local circuit flow around the site of current injection in an axon. (right panel) Local circuit flow around a propagating action potential.
The action potential normally only moves in one direction down the axon because the membrane region behind the action potential wave front is in the refractory period and unable to support a new action potential. Normally the action potential travels from the cell body down the axon towards the nerve terminal. These are called orthodromic action potentials. The axon is functionally symmetric, however, and can conduct action potentials in either direction. Action potentials propagating in the opposite direction, from the nerve terminal to the cell body, are called antidromic action potentials. If an axon is artificially stimulated in the middle of its length then an action potential can travel in two directions away from the site of current injection.
Factors Affecting the Speed of Action Potential Propagation
The speed that electrical excitation travels down an axon is much slower than the speed of current flowing through a wire. Current flow in a wire is close to instantaneous at the dimensions of typical electrical circuits, because it travels at the speed of light (3 x 108 m/s). The rate of electrical conduction in an axon is vastly slower, varying over the range 0.1 to 100 m/s.
The two major factors that affect the rate of action potential propagation are the same as those that affect the length constant; axial resistance and membrane resistance. Either increasing membrane resistance or decreasing axial resistance will result in more current flowing inside the axon, as opposed to flowing across the cell membrane. As a consequence, local circuit flow will spread over a greater length of the axon resulting in a greater spread of membrane voltage depolarization, which will initiate an action potential further down the axon and speed up the rate of conduction.
During the course of evolution two strategies have been used to increase the speed of action potential propagation:
- Decrease the resistance of the axial path down the inside of the axon by increasing the diameter of the axon, which gives a greater cross-sectional area for current flow.
- Increase the resistance of the cell membrane with a specialized sheathing, known as myelin.
Invertebrates only use the first approach and some invertebrates, such as squid, can have axons up to 1 mm in diameter. These giant axons are typically used to produce escape behavior from predators, the survival of individuals within the species being a significant factor in driving the evolutionary development of these axons. The problem with this strategy is that there is not enough space to do this with more than a handful of axons before the nervous system becomes inefficiently large. As a consequence, invertebrate nervous systems have some inherent limitations relative to vertebrates.
Vertebrates also use larger axons for the fastest conducting nerve fibers, but only up to 1 to 20 μm in diameter. The fastest vertebrate axons are faster than those of invertebrates, however, because vertebrates have developed specialized cells whose function is to increase the membrane resistance of the axon by producing a myelin sheath around the axon.
Myelination
Myelin forming cells are also known as glial cells. They form a sheath around the axon that acts somewhat analogously to the plastic insulation wrapped around the copper wire of household electrical circuits. There are periodic breaks in the myelin sheath known as nodes of Ranvier. Myelination alters the local current flow by reducing the capacitance and increasing the resistance of the internodal membrane, producing more efficient local circuit current flow. Most of the current flow across the membrane is limited to current flow through the internodal membrane.
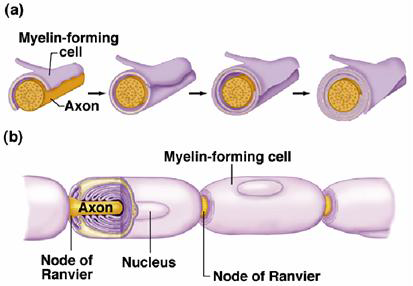
Figure 4 (a) During development myelin forming cells wrap around the axon. (b) The myelin sheath has periodic gaps known as the nodes of Ranvier.
Sodium channels are clustered at the nodes of Ranvier and are largely absent in the internodal membrane. As a consequence, a propagating action potential leaps from node to node. During action potential conduction the membrane potential in the internode region is depolarized by local circuit currents flowing from the previous node in which an action potential has fired. Conduction is fast between nodes and then slows down in the node as a new action potential is triggered (Figure 5). In effect, the action potential jumps from node to node down the axon. This is known as saltatory conduction (from the Latin ‘saltare’ meaning to jump).
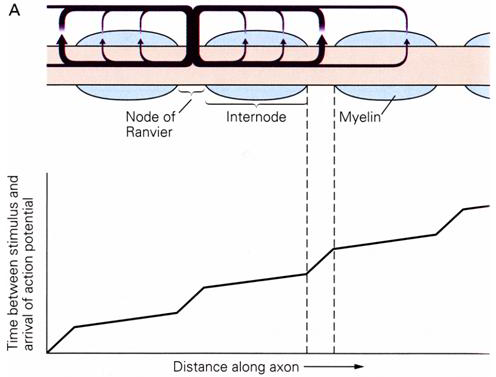
Figure 5 Excitation jumps rapidly between nodes of Ranvier.
In addition to streamlining electrical conduction, myelinated axons are also very efficient from a metabolic viewpoint. Less energy must be expended by the Na,K-ATPase in maintaining the Na+ and K+ ion concentration gradients because there are many fewer ion channels in the membrane. This is important because the axon is more susceptible to the run-down of ion gradients than the cell body, due to the much larger surface area to volume ratio of the axon.